
The Fifth Kingdom - Chapter 4a
Kingdom Eumycota
Phylum 6 - ASCOMYCOTA - the ASCOMYCETESHotlinks to: meiosporangia - life cycle
ascomata - asci - anamorphs - coelomycetes - hyphomycetes
Introduction - basic features
Zygomycetes are terrestrial fungi: there's no doubt about that. But they thrive and sporulate only in damp places where the atmosphere is more or less saturated with moisture. For example, Rhizopus stolonifer will colonize the moist interior of a loaf of bread, but won't produce its characteristic sporangiophores and mitosporangia on the outside of the bread unless the surrounding atmosphere is humid. If we persuade the fungus to sporulate by keeping the loaf in a damp chamber (a plastic bag containing a few drops of water will do) and then take it out of the bag, the sporangiophores will quickly collapse. Hyphae of most zygomycetes are wide, thin-walled, and coenocytic - continuous tubes with no cross-walls.Hyphae of the Dikarya (Phylum Ascomycota plus Phylum Basidiomycota) are narrower - although this picture shows ascomycete hyphae alongside what looks like a pine tree, it is actually a human hair. Hyphae average about 5 microns in width, but collectively they are very long (sometimes kilometres per gram of soil).

Dikaryan hyphae are also septate - they have cross-walls called septa at regular intervals. These miniature bulkheads give the hyphae some physical rigidity, and limit loss of cytoplasm if the hyphal wall is ruptured. As a result, we find that dikaryan fungi can grow in a wider range of conditions: they can often spread and fruit in drier situations than zygomycetes could tolerate.

This picture shows what you would see if you could crawl inside a hypha and look toward the septum at the end of a compartment. The septal pore is obvious, and you can also see the fine fibres of which the cross-wall is made.

Cytoplasm and nuclei can move through the septal pores of ascomycetes, as you can see in this transmission electron micrograph of a short segment of Neurospora crassa hypha - a nucleus is shown in the act of squeezing through the pore. However, Dr. Jim Aist, who has spent many hours watching nuclei in living hyphae, assures me that this does not happen very often.

Some dikaryan anamorphs (especially coelomycetes)
grow in dead leaves and stems of desert plants, and other moulds are the most drought
tolerant of all organisms, able to grow at water activities below 0.70 (for example, on
jams, salt fish and other substrates of extremely high osmotic pressure - see
Chapter
20).
While many zygomycetes can assimilate only 'accessible' substrates like sugars and starch, ascomycetes can often exploit cellulose, and many basidiomycetes can digest both cellulose and lignin, carbon sources that are available to remarkably few other organisms. Though fungi cannot fix atmospheric nitrogen (this talent seems to be restricted to the bacteria), dikaryan fungi can use many different forms of combined nitrogen: some ascomycetes even specialize in metabolizing the protein keratin, which is the main component of hair and skin. In case you were wondering if members of this group constitute a health hazard -- they do. Some other orders of ascomycetes are obligate parasites of plants. Remember the 'downy mildews' caused by oomycetes? Well, there are also plant diseases called 'powdery mildews' that are caused by ascomycetes. The similarity of terminology is unfortunate, but try to remember the difference, because although the groups of fungi involved are both obligately biotrophic, the diseases are different in many important ways, such as host ranges and methods of control. This is just one example of how taxonomy has practical implications (see Chapter 12).
Thousands of basidiomycetes, and a quite a few ascomycetes, establish intimate mutualistic symbioses (mycorrhizas) with the roots of trees, especially conifers (see Chapter 17). Nearly 18,000 ascomycetes, and a few basidiomycetes, have domesticated algae, thus becoming lichens, which can live in some of the world's harshest climates, and colonize the barest and most inhospitable substrates (see Chapter 7). Some dikaryan fungi have even re-entered the water and, lacking motile cells, have evolved other mechanisms, such as long appendages, to aid spore dispersal. Dikaryan fungi range from unspecialized, almost omnivorous saprobes, to fungi so specialized and ecologically demanding that they are found only on one particular leg of one species of insect. Some dikaryan fruit bodies are microscopic (as in many ascomycetes), but often (especially among the basidiomycetes), they are large and complex, and most of the common names applied to fungi refer to the visible teleomorphs of basidiomycetes, and in a few cases, ascomycetes. You may already be acquainted with some of these: I will introduce you to many more in the pages ahead.
Phylum ASCOMYCOTA - Characteristics of Teleomorphs
Most dikaryan fungi share a number of important features -- (1) chitinous cell walls; (2) hyphae with regular cross-walls called septa (centrally perforated to allow movement of cytoplasm, and sometimes nuclei, between compartments): (note that many yeasts are unicellular, so don't produce hyphae); (3) the ability of somatic, assimilative hyphae to fuse with one another (anastomosis) and to exchange nuclei; and (4) the occurrence in their life cycles (or at least in those which produce a teleomorph) of a unique nuclear phenomenon called the dikaryon. After sexually compatible nuclei from different mycelia have been brought together by anastomosis, they pair off, but don't fuse immediately to form a diploid zygote. Instead, they go on dividing synchronously to populate what are called dikaryotic hyphae, in which each compartment has two sexually compatible haploid nuclei. Oh yes, they do fuse eventually, but not before some remarkable developments have taken place, and in basidiomycetes, perhaps not for years. Read on.
If ascomycetes and basidiomycetes share all these things, how do they differ? Actually, in many ways, and with experience it's usually easy to tell their sexual fructifications apart with the naked eye. But their microscopic, unicellular meiosporangia are most diagnostic of all (compare the two sets of diagrams below).
Upper diagrams - the meiosporangia of ascomycetes are
asci
(singular, ascus). They are cylindrical or sac-like and at maturity
usually contain eight haploid spores (ascospores) which
are expelled into the air through the top of the ascus.

Lower diagrams - the meiosporangia of basidiomycetes are basidia
(singular, basidium): they usually have four tiny
projections called sterigmata, each bearing a haploid spore (basidiospore)
which is shot away individually at maturity. The formation of asci or basidia marks the
end of the dikaryophase: the paired nuclei have fused and the resulting zygote has
undergone meiosis (and a mitosis in ascomycetes) to produce 8 haploid ascospores or 4
haploid basidiospores. By comparing the two sets of diagrams above, note how
similar the developmental processes are until the final stages.
Teleomorph life cycle
Now let's examine the sexual (teleomorphic) part of the ascomycete life
cycle from the beginning (bottom). When an ascospore germinates, it establishes a haploid
mycelium. In heterothallic ascomycetes, this can't undergo sexual reproduction until it
meets another compatible haploid mycelium.

When this rare event
takes place, the fungus cleverly maximizes the ensuing potential for genetic
recombination. One would expect a single sexual fusion, resulting in a
single zygote.
But most ascomycetes interpolate a dikaryophase, during which the number
of pairs of compatible nuclei is multiplied, often enormously, as dikaryotic hyphae (often
called ascogenous hyphae, as in the diagram above) grow and branch within a mass of
monokaryotic (haploid) tissue which is the framework of the fruit body (the ascoma).
Eventually, the ultimate branches of the dikaryotic hyphae, of which there may be
millions in larger ascomata, reach their ordained positions in the future hymenium and the
long-delayed sexual fusions take place. The genome
is reshuffled during the ensuing meiosis in each ascus (this genetic recombination is due
to crossing-over, which is
explained in Chapter 10)Each meiosis will produce a somewhat different
arrangement of the genome. In this way the products of a single encounter are first
multiplied, then meiosis generates a lot of genetic diversity. Not only is the dikaryon
itself an unusual phenomenon, but during the dikaryophase an effectively diploid mycelium
is growing within, and drawing nourishment from, the haploid ascoma tissue. This
phenomenon has interesting parallels in the red algae, though molecular evidence doesn't
support a close relationship between the two groups (both lack motile gametes and appear
to have simply hit upon the same solution to the problem posed by the rarity of sexual
encounters).
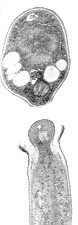
Ascospores are not motile, in the sense of self-propelling, but most ascomycetes nevertheless send their ascospores on their way with a burst of kinetic energy. The ascus is a tiny spore-gun, which works by building up internal pressure, then releasing it through the tip (see animation below). The job of most asci is to get their ascospores into the turbulent airflow above the ascoma. Before the ascus bursts open at the tip and shoots its spores skywards, the pressure inside the ascus is cranked up by osmolytes such as glycerol. It builds up to about 3 atmospheres before the apex blows.
Mature asci of the dung-inhabiting Ascobolus
(below) project above the hymenium and point toward the light before discharging their
spores. In this way they ensure that the spores will not run into any obstacles on their
upward flight (see Chapter 8).
Four kinds of Ascoma.
The multicellular structures (ascomata) that produce the asci, and act as the platforms
from which the spores are launched, come in four main designs, sectional views of which
are shown in the diagrams below.
(1)
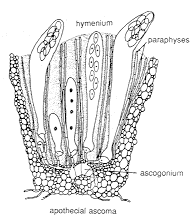
APOTHECIAL ASCOMATA [below]
The construction of the ascoma may allow several or many
asci to discharge simultaneously because the entire fertile layer or hymenium is
exposed...
The diagram on the left shows a vertical section through a very small
apothecial ascoma.
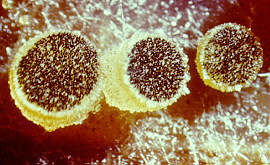
The photomicrograph on the right shows three ascomata of
Ascobolus:
the dark dots are unitunicate-operculate asci (see next section)
containing dark spores, arranged in a flat superficial hymenium.
...or it may permit discharge of only one ascus, or a few asci, at a time, as in
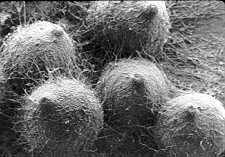
(2) PERITHECIAL ASCOMATA
[below] which contain
unitunicate-inoperculate asci, (see the next section of the chapter).
The left hand scanning electron micrograph shows what many perithecial
ascomata look like. The diagram in the middle is of a vertical
section through a perithecial ascoma and its neck. The right hand
photomicrograph is of a translucent ascoma inside which the asci, with
their rows of dark ascospores, can be seen.


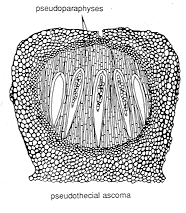
and (3) PSEUDOTHECIAL ASCOMATA [below], which contain
bitunicate asci (see the next section of the chapter)...

...or they may lack an opening entirely
in
(4)
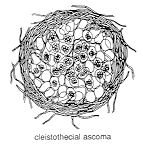
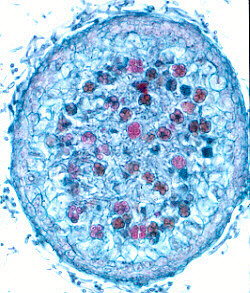
CLEISTOTHECIAL ASCOMATA(below).
The asci in
these are often spherical, as in the illustrations below, and no
longer shoot their spores: the fungus has evolved a new dispersal strategy. That may
have happened because the fungus fruits in a confined space (for example, under bark, or
below the surface of the ground) where airborne dispersal cannot operate. We often find
that the spores of such fungi are dispersed by animals. Both diagram
and photomicrograph show sections through a cleistothecial ascoma: the
globose asci are clearly visible, and it is equally clear that they are
not arranged in a layer or hymenium, as they were in the other three
kinds of ascoma.
Four kinds of
Asci. Before we go on to explore the many orders of
ascomycetes, we must take a closer look at the ascus itself. All asci are not the same.
There are four flavours...
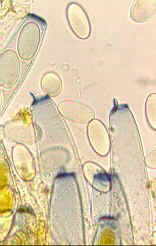
(1) Unitunicate-operculate
asci
Unitunicate asci have a single wall. Some
have a built-in lid or operculum (stained blue - bottom left) - at maturity this pops open
around a built-in line of weakness (below, right) so
that the spores can be ejected.
Unitunicate- operculate asci are found
only in apothecial ascomata. Some of the operculate asci in the colour picture
(middle) have discharged their spores; one has not.
The animation on the right (courtesy of Dr. Jim
Worrall) shows
how
operculate asci develop and shoot.

![]()
(2) Unitunicate-inoperculate
asci
Other asci have no operculum, but have a special elastic ring mechanism
built into their tip. This is a pre-set pressure release valve, or sphincter, and the ring
eventually stretches momentarily, or turns inside out, to let the spores shoot through.
Such inoperculate asci are found in
perithecial and some apothecial ascomata.
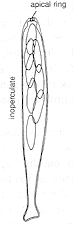


The two colour photomicrographs (above, centre and right) are of the same
asci, only
one of which is mature. The centre picture was taken by bright-field
illumination, and shows that the apical rings have been stained blue by iodine: you can
hardly see the ascus walls. The right-hand picture is by phase contrast, and shows
the unstained asci more clearly.
The transmission electron micrograph (below - courtesy of A. Beckett) shows
a section directly through an ascus tip of Xylaria. The apical apparatus is
actually shaped like a ring, a doughnut or a torus, depending on your background.

(3)
Prototunicate asci have no active spore-shooting mechanism. These
asci are usually more or less spherical, and are found in cleistothecial (occasionally
perithecial), and hypogeous ascomata. Sometimes the wall of this kind of ascus dissolves at maturity and
releases the ascospores, which can then ooze, rather than be shot, out of the ascoma; or
they may wait inside until it decays or is ruptured. These asci are often called prototunicate.
Yet perhaps because they are found in several otherwise rather different orders, it seems
likely that they represent a secondary condition, and have evolved several times
from unitunicate asci (as they clearly did in the truffles - Tuberaceae
in Chapter 4b). The TEM photo is of a very thin section of a single ascus of Eleutherascus,
which passed through only 4 of the 8 spores.


4) Bitunicate asci
have a double wall.
A thin, inextensible outer wall covers a thick, elastic
inner wall. At maturity the thin outer wall splits, and the thick inner wall absorbs water
and expands upward, carrying the ascospores with it (bottom
left). This 'Jack-in-a-box' design
allows the ascus to stretch up into the neck of the ascoma to expel its
spores. The
bitunicate ascus is so different from the unitunicate ascus that we assume they diverged a
long time ago. Note that the ascospores in the colour photomicrograph (bottom right) are
darkly pigmented dictyospores,
with septa running both across and along the spore).


In many unitunicate ascomycetes, the perithecial ascoma develops only after the sexual
stimulus, so that the asci can grow into an actively enlarging cavity. In many bitunicate
ascomycetes, fertilization doesn't happen until after a solid primordium or stroma has
developed, so room has to be made for the asci by dissolving away existing tissue. In some
cases the asci themselves do the job, but in others it is carried out by special sterile
hyphae (pseudoparaphyses) growing down from the upper layer of the stroma; the asci then
grow up between them. Remember that pseudothecial ascomata always produce
bitunicate asci.
Subphylum ASCOMYCOTINA -
Many Kinds of Anamorph
Here is a mantra to begin with - say it until you know it:
In any modern consideration of the
ascomycetes, we cannot ignore
their asexual reproductive phases, many of which are called moulds. You already know that
zygomycetes have diverse asexual phases. So you won't be surprised to discover that many
ascomycetes have comparable asexual (anamorphic) phases during which they reproduce
rapidly, and often relatively cheaply, by means of mitospores called conidia.
The asexually reproducing phase of the ascomycete life cycle was more or less ignored for
many years in favour of teleomorph studies. But when we consider that the anamorph is an
important (and sometimes the only) phenotypic expression of many ascomycete genotypes, we
realize that it has much to tell us. Besides, one can get DNA and RNA from
anamorphs just as easily as from teleomorphs, so we are beginning to understand the
relationships of anamorphs better, even in many cases where no teleomorph is known.
Though they play essentially the same role in the life cycle, the anamorphs of
ascomycetes differ from those of zygomycetes in two very important respects:
(A) While zygomycete mitospores commonly originate by free-cell formation
inside a sporangium, many spores cleaving from a single mass of cytoplasm, the mitospores
(conidia) of ascomycetes are basically modified bits of hyphae, either budded out as a new
structure, or converted from a whole existing cell.
(B) In zygomycetes, anamorph and teleomorph often occur together (especially in
homothallic species) and always share the same binomial. In ascomycetes, anamorph and
teleomorph often develop at different times, and on different substrates. Each phase has
often been collected in total ignorance of the existence of the other, and because of
this, the International Code of Botanical Nomenclature maintains that it is legal to give
them separate binomials - a useful option but one giving rise to a great
deal of confusion in some students' minds (How can one organism have two
names?)
Even though several thousand anamorph-teleomorph connections have now been established in
the ascomycetes (you can now find them at this web site:
http://www.wi.knaw.nl/collections/
)
these represent only a small proportion of the total number of taxa,
either of anamorphs or teleomorphs (we know about 30,000 of each). Because anamorphs so
often occur alone, it is still normal and accepted practice to use separate binomials for
them, as you will see. Nevertheless, I find it completely unacceptable to talk about
conidial fungi, as many other texts do, as if they constituted a separate major high-level
taxon called the 'Deuteromycotina.' This ignores both the evidence that they are all
expressions of dikaryan genomes, and the thousands of connections that have already
been established with teleomorphs (and the number grows every year).
Of about 30,000 known ascomycetes, about 5,000 have so far been connected to their anamorphs. What about the many thousands of conidial (anamorphic) fungi that are still 'orphans'? I think there is good reason to believe that many of them have given up sex altogether, and become 'anamorphic-holomorphs,' though they seem to have retained some genetic flexibility by: (a) having more than one kind of nucleus in their mycelia (heterokaryosis) as a result of occasional hyphal fusions (anastomoses); (b) sometimes undergoing a complex parasexual process involving rare somatic diploidization, mitotic crossing-over, and finally a return to the haploid condition (this process is more fully explained in Chapter 10).
It turns out that the conidial fungi are a mixed bag. Although most of them are
(or were) part of ascomycete life cycles, some are (or were) connected
with basidiomycetes. Despite this mixed ancestry, we have had to set up a
single classification for all of them, because it is often impossible to tell in which
subphylum the connection lies. Unfortunately, our scheme for classifying the anamorphs has
so far been able to make little reference to teleomorphs, for several reasons:
(1) teleomorphs are known for only 10-15% of anamorphic species;
(2) members of what seems to be a single anamorph genus may have teleomorphs in many
different holomorph genera (even from different orders). This must be due to convergent
evolution among anamorphs;
(3) anamorphs belonging to several different anamorph genera can have sexual phases in a
single holomorph genus. This must be due to radiative evolution among anamorphs.
So our attempts to classify anamorphs have concentrated on:
(A) their mitospores (conidia),
(B) the diverse structures (conidiogenous
cells, conidiophores and conidiomata) which bear them, and
(C) the ways in which they develop (conidiogenesis).
Molecular techniques are now helping us to elucidate their relationships.
Morphology of Anamorphs
There are about
1,500 described
genera of conidial fungi, and almost 30,000 described species, and these numbers are
increasing rapidly. The first really useful classification of dikaryan anamorphs,
established in the late 19th century, was based on mature morphology. The principal
characteristics used were:
(1) colour, septation and shape of conidia;
(2) conidiophore aggregation, or lack of it;
(3) the production of conidia in enclosed structures or the absence of such enclosure.
Don't worry, all these characters are illustrated below.
The division of anamorphs into two large groups is informal, is based on morphology, and
is really just for convenience.
The easiest decision to make is usually whether a conidial fungus is a hyphomycete or a coelomycete. You can recognize a hyphomycete because its conidiophores can be single or aggregated in various ways, but are never enclosed within a covered conidioma. Coelomycetes form their conidia in enclosed conidiomata (below), which usually develop just beneath the surface of their plant substrate. I am not going to discuss coelomycetes in detail (though there are thousands of them, and they cause many important diseases of crop plants), but I must mention a few basic facts.
Coelomycetes = covered
conidiomata
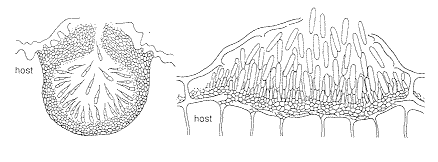
The first kind of enclosed conidioma is the acervular conidioma
or acervulus [above, right]. This may develop at various depths within the host: it can be
subcuticular (covered only by the host cuticle); intraepidermal (arising within the cells
of the epidermis, as in the diagram); subepidermal, as in the photomicrograph below; or
developing beneath several layers of host cells.
Under this roof of host material, fungal hyphae aggregate and
form a flat fertile layer of short conidiophores that produce many conidia (here shown in
a vertical section).
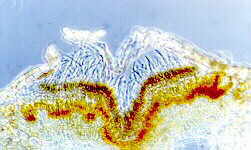
The pressure of accumulating conidia, and often of accessory mucilage, eventually splits the host epidermis and allows the conidia to escape. These acervuli have developed in the surface layers of an avocado.

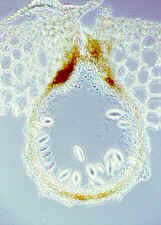
At the other end of the spectrum is the flask-shaped pycnidial conidioma or pycnidium, seen from above (far left), and in vertical section (bottom, right), in which the fungus itself provides the enclosing wall, and conidia eventually ooze out through a narrow apical ostiole.

'Mostly Moulds'
Hyphomycetes = exposed conidiophores or conidiomata
Most of what we call moulds
are hyphomycetes (the anamorphic or asexual reproductive phases of many Ascomycetes and
some Basidiomycetes). As of May 2005, we
recognize almost 1,400 form-genera and almost 13,000 species of
hyphomycetes.
A small minority of moulds are anamorphic
phases of Zygomycetes (things like Rhizopus and Mucor),
or microscopic ascomycetes such as Chaetomium. Weeds are plants
growing where we don't want them. Moulds are sometimes the weeds of the
fungal world -- fungi which have a habit of fruiting on substrates that
are of interest to us, such as food, paper, leather and wood.
(1) container for making a shape; root 12th century, old French 'modle'.
(2) soil rich in humus; root old English 'molde'.
(3) fungi involved in food spoilage and some kinds of decay; root, 15th century English 'moul', to go mouldy.
Note that the last root is the only one containing a 'u'. Ergo, the spelling 'mould' clearly and correctly differentiates the fungus from the other two].

But put a bit of that black stuff under the microscope, and a new world opens up... Among the hyphomycetes, conidiophores are usually solitary [below, left], though they sometimes form columnar aggregations called synnematal conidiomata [below, centre], or cushion-shaped masses called sporodochial conidiomata [below, right].

On the left are two individual conidiophores of a darkly pigmented hyphomycete called Periconia. On the right, a synnematal conidioma of Gangliostilbe
(a number of conidiophores growing up in a parallel bundle)

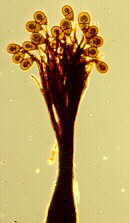
Here is a photo of sporodochial conidiomata of Trimmatostroma salicis.
I found this fruiting on twigs of a weeping willow in our garden.

Here are the conidia produced in those sporodochia. How shall we classify them?

Well, now come the morphological characters of the microscopic conidia (asexual spores).
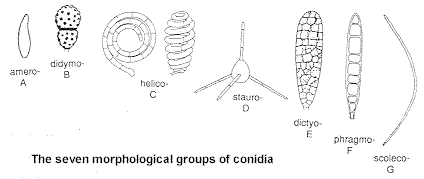
Presence or absence of pigment is important, as are shape and septation.
There are seven s & s categories, all illustrated above.
(B) the addition of one cross-wall makes them didymospores.
(C) conidia curved through more than a half-circle, or coiled in two or three dimensions, are called helicospores.
(D) those with several conspicuous, radiating arms or other projections are called staurospores.
(E) septa running two ways, like the meshes of a net, or like the mortar layers of a brick wall, identify dictyospores.
(F) two or more transverse septa, arranged like the rungs of a ladder, characterize phragmospores.
(G) finally, those which are long and thin (more than 15 times as long as they are wide), are called scolecospores (which means 'worm-like'). Practise by putting names on the photos below...





Can you name the kinds of conidia illustrated in these five photomicrographs? It's easy if you refer back to the diagrams given above.
The Trimmatostroma
conidia shown above are mostly phragmospores, but some appear to be
dictyospores, so we would list both in our description of the fungus.
You might think that all coiled conidia (helicospores)
would be truly 'related', but check out the subsidiary page on what
molecular studies can tell us about relationships among fungi producing
helicospores.
Conidiogenesis (spore development)
More recently, it was
discovered that conidial fungi use a number of different techniques to produce their
spores. Since these represent genuine `embryological' differences, they have become
important characters in our classification. Spores which look alike often develop in
different ways. We begin by checking an anamorph to see which of two basic patterns
of development - blastic or
thallic - it exhibits.
In blastic conidiogenesis [A, below,
upper left], the young conidium is
recognizable before it is cut off by a cross-wall (this is an extension of the idea of
cells `budding').
In thallic conidiogenesis [B, below,
upper right], the cross-wall is laid down
before differentiation of the conidium begins.

Ripe conidia may also be liberated in two basic ways,
schizolytic and rhexolytic. In
schizolytic
dehiscence (C, above, left), the halves of a double septum split apart by the breakdown of a
kind of middle lamella. In rhexolytic dehiscence (D,
above, right), the outer wall of a
cell beneath or between conidia breaks down.
We will examine eight different kinds of conidium development: six are blastic, two thallic.
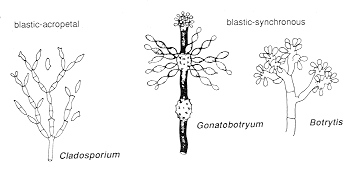
Type I - blastic-acropetal or blastic-synchronous conidiogenesis
The Monilia anamorph of Monilinia fructicola
(Unitunicatae-Inoperculatae: Leotiales), the brown rot fungus of peach and other stone
fruits (see Chapter 12), and the
Cladosporium anamorph of Mycosphaerella tassiana (Bitunicatae:
Dothideales),
a common mould on decaying organic matter; produce conidia in chains by apical budding.
The youngest conidium is at the tip of the chain. The chain branches when two buds, rather
than one, develop on a terminal conidium (which may then be called a ramoconidium).


The animation
(left) shows how acropetal chains of conidia develop and branch. This is clearly just a modified form of hyphal growth.
The hyphomycetous anamorphs Botrytis
and Gonatobotryum produce many conidia synchronously on a swollen cell: Gonatobotryum
(above) goes
on to form acropetal chains of secondary conidia, while Botrytis
(two SEMs of
successive stages, below) does not. Below
you can see how Botrytis deals with a pumpkin after Hallowe'en: the
grey patches are made up of thousands of conidiophores producing millions of
conidia.

Botryosporium (below, left) also produces conidia
synchronously on swollen cells. In this genus, the branches
bearing these vesicles are arranged along an extremely tall, graceful,
white conidiophore up to 2mm long (below, left) in a sequence from
youngest at the tip to oldest near the base. So in this fungus, you can often see all stages of conidiogenesis on a
single conidiophore (below, right). This fungus, which often turns up in greenhouses growing on dead
leaves, is sometimes called 'the beautiful hyphomycete.' (Pictures
courtesy of Dr. Roland Weber)


The Chromelosporium anamorph of Peziza has a simpler
conidiophore, with longer vesicles.
The sequence of blastic-synchronous conidium development is shown in the three accompanying photomicrographs...
(the first two are taken by Nomarski interference contrast, the
third by phase contrast)



Type II - blastic-sympodial conidiogenesis
In species of Beauveria, hyphomycetous insect
pathogens which are now being used in biological control of potato beetle, the narrow apex
of the conidiogenous cell extends sympodially: each new apex becomes converted into a
blastic conidium, then the next apex grows out from behind and to one side of it. The more
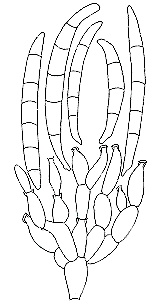
conidia are produced, the longer the conidiogenous cell becomes. Although Leptographium
anamorphs of Ophiostoma (Prototunicatae: Ophiostomatales) have single
conidiophores, these have complex heads with several tiers of supporting cells
(metulae),
the ultimate ones bearing many sympodially (or percurrently) extending conidiogenous
cells, and innumerable conidia accumulate in a slimy head; these spores are
insect-dispersed. Basifimbria (teleomorph unknown), which is common on horse
dung, has simple conidiophores that elongate sympodially during conidiation.

The sequence of time-lapse photos (below, left) shows how
conidiogenous cells
elongate sympodially with repeated conidium formation. The SEM photos (below,
right) show how a
conidiogenous cell becomes geniculate (bent like a knee) as conidia form,
and how scars are left as
they secede (pictures from Cole and Samson 1979)


Under the microscope, these dark spots are shown to bear a halo of sympodially extending conidiophores, which have a rather unusual regular left-right-left-right alternation of the extensions (below, left). These conidiophores bear obpyriform didymospores (below, right)


Here is another fungus that has sympodial conidiogenesis - Pleiochaeta setosa. Note the repeatedly geniculate conidiogenous cells (and the pigmented phragmosporous conidia with apical appendages -- lots of good characters here!).

Type III - blastic-annellidic or blastic-percurrent conidiogenesis
In the Spilocaea anamorph [below] of Venturia
inaequalis, the apple scab fungus, each seceding conidium leaves a ring-like scar, an
annellation, around the conidiogenous cell, which then grows on through the scar
('percurrently') to produce the next conidium. Conidiogenous cells that have produced x
spores bear x annular scars -- hence the name annellidic.
The photo below is an oil-immersion picture of a tape-lift from one of my
apples (which is unfortunately not resistant to scab). You can see
flame-shaped conidia of Spilocaea (left and right) with truncate bases, and several annellations on the
central conidiogenous cell, which is just developing a new conidium

The photo of Annellophora africana (below) shows many, widely-spaced annellations, each of which was the level at which a conidium was formed and released.

The time-lapse
sequence of Scopulariopsis (below) shows how annellations can also be extremely inconspicuous and closely spaced. If you look
carefully, you can see that the conidiogenous cell is only slightly longer after producing
seven conidia.

It has recently been confirmed that some individual anamorphs can be both annellidic
and sympodial. I'll give you two examples of how this knowledge may change our
classification. When it was thought, not many years ago, that conidiogenesis in the
synnematal anamorphs of Ophiostoma species was either exclusively sympodial or
exclusively percurrent, they were segregated into two anamorph genera (Pesotum
and Graphium). Now it has been shown that both kinds of conidiogenesis can
occur on the same conidiophore, they are being united again in the older genus, Graphium.
In exactly the same way, some complex mononematous anamorphs of Ophiostoma
species were segregated into the 'exclusively sympodial' Verticicladiella and the
`exclusively percurrent' Leptographium, but are now united under the older name, Leptographium.
Type IV - blastic-phialidic conidiogenesis
Many common moulds produce conidia in rapid basipetal succession from the open end of special conidiogenous cells called phialides. Important genera such as Penicillium, Aspergillus, Fusarium, Stachybotrys, Trichoderma and Chalara are all phialidic.


This picture is of a glass model of Aspergillus herbariorum, made
in 1929 by R. Blaschka, and part of the wonderful Ware collection of glass sculptures of
fungi at Harvard University Botanical Museum.

Below is Trichoderma harzianum, a well-known mycoparasite.

Many plant pathogenic hyphomycetes, such as Fusarium [below left] and Verticillium
[below right], both causing serious wilt diseases of crop plants, produce phialides.

Most phialides don't change in length while producing many successive conidia, though
many wall layers build up inside the open end of the cell (TEMicrograph, below, and
diagrams).
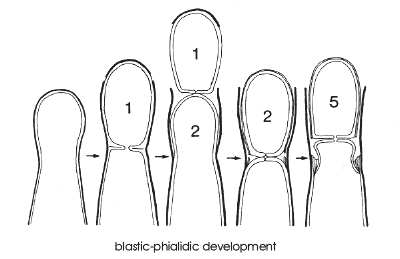
This accumulation of wall layers may eventually plug the opening, and in phialides
to which this happens there is a tendency to produce sympodial
extensions that develop new fertile apertures. Such phialides are called polyphialides
since they have more than one conidiogenous locus.
Penicillium and Aspergillus are dry-spored, Fusarium, Verticillium
and Stachybotrys have slimy spores. Phialidic ontogeny is basically rather
similar to type III - percurrent.
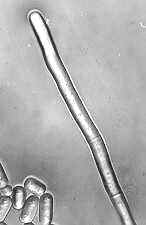
Acremonium is morphologically one of the simplest phialidic anamorphs -
a single tapering colourless phialide with a small droplet of slimy
colourless amerospores accumulating at its tip. Yet this apparent
simplicity is belied by the fact that holomorphs of Acremonium are
found in no fewer than 40 ascomycete genera, distributed among the
Nectriaceae, Cephalothecaceae and Chaetomiaceae. It has
also been found that species of Acremonium are allied to more complex
anamorphs: A. strictum with Sarocladium (rather penicillate),
A. crotocinigenum with Trichothecium (branching conidiophores,
blastic retrogressive conidiogenesis, didymospores) and A.
cucurbitacearum with Plectosporium (didymospores).
And of course, the Acremonium-like anamorph of Epichloë
(Clavicipitaceae) is now called Neotyphodium.
Caveat spectator!
Type V - blastic-retrogressive conidiogenesis
In Basipetospora (a thermotolerant fungus used in Indonesia in the preparation
of a red food colouring), a conidium forms at the tip of the relatively undifferentiated
conidiogenous hypha and is delimited by a cross-wall; then a short zone of the hypha just
below the conidium balloons out to produce the second conidium. After this has been
delimited by a septum, the next segment of the hypha plasticizes and blows out, and so on.
As the chain of conidia elongates, the conidiogenous hypha becomes shorter.
Watch the animation several times. Focus first on the elongation of the chain, then on
the shortening of the conidiogenous hypha. Similar development occurs in Trichothecium
and Cladobotryum, but this is an unusual kind of conidiogenesis.


Type VI - basauxic conidiogenesis

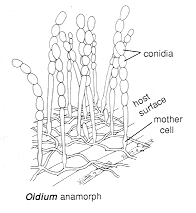 In
In
the Oidium
anamorph of Erysiphe graminis, whitish chains of conidia (the
'powdery mildew')
cover the host leaves. Each chain consists of a graded series of gradually maturing
conidia, the oldest at the tip, the youngest barely differentiated from the hyphal cell
just below it. New material is added at the base of the chain in a form of intercalary
growth, arising from a sometimes swollen mother cell which appears to be a highly modified
phialide.
Type VII - thallic-arthric conidiogenesis
In
the Geotrichum anamorphs of Dipodascus spp. (Saccharomycetes) (right),
an assimilative hypha stops growing, then becomes divided up into short lengths by
irregularly arising septa. These are double septa which split apart schizolytically to
give a 'chain' of short cylindrical 'fission arthroconidia' that disarticulates and
appears jointed (hence 'arthric').


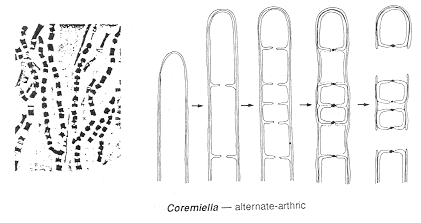
In Coremiella some hyphal cells degenerate to release the
intervening cells as 'alternate arthroconidia.'
In Oidiodendron, a common soil mould, the branches of an often
tree-like conidiophore disarticulate into conidia, ultimately leaving only the denuded
'trunk,' (the stipe).

Many basidiomycetes also produce thallic-arthric conidia.

Type VIII - thallic-solitary conidiogenesis
The Microsporum
anamorphs of Nannizzia (Prototunicatae: Onygenales), which can digest keratin,
and cause skin diseases in humans (see Chapter 23), develop large thallic
phragmospores at the ends of hyphae. These conidia are liberated rhexolytically.

Here are some fine phase-contrast photomicrographs of thallic
conidiogenesis from Dr. J.W. Carmichael. The picture at top left is of
phragmosporous thallic-solitary conidia of Microsporum, that at
top right of alternate arthric conidia of Trichophyton, that at
bottom left of Coremiella (alternate arthric); that at
bottom right of Chrysosporium (also alternate arthric).

Finally, on the conidiogenesis front, here is a
diagram modified from an excellent book by Cole and Samson (1979 - see
references at end of Chapter 4b) with labels identifying no fewer
than 12 variations on the developmental theme. Note those not
discussed above, and pursue the matter in the original book, if you are
keen...

One of the earliest kinds of
'developmental' information was the observation that some
moulds produced conidia in chains, while others did not. But looking back over the various
kinds of conidiogenesis I have just described, I think you could probably come up with no
fewer than seven ways in which look-alike 'chains' of conidia can develop. These are seven
good reasons why we no longer rely on mature morphology in establishing our classification
of conidial fungi. Now there's a good exam
question...
Importance of Conidial Anamorphs
Why is it important to be able to identify conidial fungi? Some of them literally grow on you. The most prevalent fungal diseases of humans (mycoses) are caused by conidial fungi. Various superficial mycoses, ranging from ringworm of the scalp, through jock itch (more vulgarly known as crotch-rot), to athlete's foot, are caused by keratin-attacking species of Microsporum, Epidermophyton, and Trichophyton (see Chapter 23).
When I compiled the fungi causing important plant diseases, I found that 62 were
conidial fungi (hyphomycetes and coelomycetes), as compared to 111 from all other fungal
groups combined. One of the most serious outbreaks of plant disease in recent years was
the southern corn blight, caused by Bipolaris (Helminthosporium) maydis, the
anamorph of Cochliobolus heterostrophus (Bitunicatae: Dothideales), which
devastated the U.S. corn crop in 1970. The special 'Texas male sterile' strain of corn
becoming widely used for seed at that time was highly susceptible to the fungus, which
produces a toxin that disrupts membranes, especially those of the mitochondria, reducing
the production of ATP. The toxin also reduces photosynthesis: it inhibits uptake of
potassium by the guard cells of the stomates, causing the stomates to close and thereby
reducing the intake of carbon dioxide. Southern corn blight was brought under control by
changing the strain of seed corn used by growers. Plant diseases are covered in more
detail in Chapter 12.
Cellulolytic hyphomycetes cause blue stain and soft rot of wood,
discolouration and loss of strength of cotton materials (the phialidic Stachybotrys
chartarum shown here is particularly troublesome in the tropics), and moulding of almost any damp
organic substrate. Stachybotrys also commonly grows on
paper (such as that covering gypsum wallboard) in damp
buildings, and is now regarded in some quarters as a serious threat to
human health, though there is no convincing scientific evidence for this
assumption.

Here, colonies of Penicillium italicum
are shown growing on oranges: this exemplifies the moulds that spoil food in storage
(e.g., in your refrigerator). Food spoilage is the subject of Chapter
21

Worse yet, Aspergillus flavus (of which a single highly magnified phialide-bearing conidiophore is shown here), grows on peanuts and many other substrates, producing a mycotoxin called aflatoxin, which contaminates food and causes liver damage even at very low concentrations. It is the most potent carcinogen (cancer-inducing) substance known.
The subject of mycotoxins is explored more fully in Chapter 21.

The 'genome sequences' or genetic maps for the fungi Aspergillus
fumigatus, Aspergillus nidulans and Aspergillus oryzae
have now been decoded. Although these three species are from the
same genus, they have been found to be as genetically different as
fish and man (groups which diverged 450 million years ago), since
they share only about 68% of the same proteins. They also differ
considerably in genome size, with that of Aspergillus oryzae
being 31% bigger than that of Aspergillus fumigatus and 24%
bigger than that of Aspergillus nidulans. Intriguingly, over
30% of the 9,500-14,000 genes identified are new to science and of
unknown function. Aspergillus fumigatus was identified as a
cause of infection as long ago as 1848 and is now the leading
infectious cause of death in vulnerable leukaemia and bone marrow
transplant patients (see
Chapter 23).
Aspergillus nidulans has been a leading experimental genetic
system for the last 50 years, whilst Aspergillus oryzae has
been used in the Far East for 2,000 years to produce sake (rice
wine), miso (soybean paste) and shoyu (soy sauce) (see
Chapter 19).
Fusarium graminearum, growing on feed
corn, produces another mycotoxin, zearalenone, which is a steroid, and
causes oestrogenic syndrome - vaginal and rectal prolapse - in young
female pigs. Many other mycotoxins have been discovered in recent years. They are
potential threats to human and animal health of which we are only now becoming fully
aware, and they have necessitated the development of new techniques for toxin monitoring
and new programs for plant protection and food storage.
On the positive side, hyphomycetous and coelomycetous anamorphs are among the prime colonizers and decomposers of plant debris, playing a vital role in the carbon and nitrogen cycles. Hyphomycetes dominate the soil mycota in most forests. The economy of many streams is based on the dead leaves of land-based plants. These are colonized by aquatic hyphomycetes, which usually form tetraradiate (four-armed) conidia, and are tolerant of low temperatures so can grow during the winter and even under ice. These fungi make the dead leaves much more palatable and nutritious for the various detritivorous invertebrates which eat them, and thus the fungi act as vital intermediaries in energy flow in northern stream systems. The terrestrial and aquatic ecology of conidial fungi is explored in Chapter 11.
Some soil-inhabiting hyphomycetous anamorphs have evolved specialized traps with which they catch small animals -- nematodes, rotifers, tardigrades, amoebae and even springtails -- these truly predatory fungi can be visited in Chapter 15.
Conidial fungi are not just out there doing their own thing. We have also learned to exploit some of them for our own purposes. The enzymes of Penicillium camembertii produce the soft, smooth texture of Camembert and Brie cheeses. Penicillium roquefortii puts that inimitable zippy flavour in blue cheeses such as Roquefort, Danish Blue, Stilton and Gorgonzola. Now there's even Cambozola, which blends the buttery texture of Camembert with the assertive flavour of Gorgonzola.
Aspergillus oryzae is used in the Far East to turn soya protein into such delicacies as soy sauce (or its sweet Indonesian variant, Ket-jap, the word which became Ketchup in English)(see Chapter 19). Other anamorphs are also exploited in the orient to prepare a variety of "fermented foods."
Finally, despite the insidious threat of mycotoxins, other secondary metabolites have
come to play important roles on the human stage: (1) a substance which gave Penicillium
chrysogenum an edge over competing bacteria in the natural habitat became one of our
most potent weapons against bacterial disease - penicillin.
(2) Cyclosporine, a metabolite isolated from the
mould, Tolypocladium niveum (above), is the most effective and least toxic
immunosuppressant yet discovered.
It has enormously improved the success rate of organ transplant operations by
preventing recipients' immune systems from rejecting the implant, but not leaving them
totally defenceless against infection. This substance or its derivatives also hold
out some hope for treatment of severe auto-immune diseases such as juvenile diabetes,
rheumatoid arthritis, multiple sclerosis, myasthenia gravis, aplastic anaemia, Addison's
disease, Hashimoto's thyroiditis, systemic lupus erythematosus and
Henoch-Schonlein purpurea.

In addition, conidial anamorphs can now be genetically transformed to act as hosts for
vectors carrying multiple copies of genes from other eukaryotic organisms (including
humans), and have already been persuaded to express and secrete a number of eukaryotic
gene products, including insulin, human growth factor, human tissue plasminogen activator
(used to dissolve blood clots), bovine chymosin (an enzyme used in cheese-making), and
amylase and cellulase enzymes (see Chapters 10, 24). Obviously, the
biotechnological potential of the moulds is tremendous. The second part of
this chapter (4b) examines the various orders of Ascomycetes...the
holomorphs.

Here is a quiz which will tell you how well you have
retained the information about development of conidia. See how many of the 12
methods you can recognize by name.
Make a list, then check it by clicking here.
Go to Chapter 4b
Go to Table of Contents
© Mycologue publications 2020